Baryonyx
| Baryonyx Temporal range: Early Cretaceous, 130–125 Ma | |
|---|---|
 | |
| Reconstructed skeleton, Natural History Museum, London | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Family: | †Spinosauridae |
| Subfamily: | †Baryonychinae |
| Genus: | †Baryonyx Charig & Milner, 1986 |
| Species: | †B. walkeri |
| Binomial name | |
| Baryonyx walkeri Charig & Milner, 1986 | |
Baryonyx (/ˌbæriˈɒnᵻks/) is a genus of theropod dinosaur which lived in the Barremian stage of the early Cretaceous Period, about 130–125 million years ago. The holotype specimen was discovered in 1983 in Surrey, England, and the animal was named Baryonyx walkeri in 1986. The genus name, Baryonyx, means "heavy claw" and alludes to the animal's very large claw on the first finger; the specific name (walkeri) refers to its discoverer, amateur fossil hunter William J. Walker. Fragmentary specimens were later discovered in other parts of the United Kingdom and Iberia. The holotype specimen is one of the most complete theropod skeletons from the UK, and its discovery attracted media attention.
Baryonyx was about 7.5 m (25 ft) long and weighed 1.2 t (1.3 short tons), but the holotype specimen may not have been fully grown. It had a long, low snout and narrow jaws, which have been compared to those of a gharial. The tip of the snout expanded to the sides in the shape of a rosette. Behind this, the upper jaw had a notch which fitted into the lower jaw (which curved upwards in the same area). It had a triangular crest on the top of its nasal bones. Baryonyx had many finely serrated, conical teeth, with the largest teeth in front. The neck was less curved than that of other theropods, and the neural spines of its dorsal vertebrae increased in height from front to back. It had robust forelimbs, with the eponymous first-finger claw measuring about 31 cm (12 in) long.
Now recognised as a member of the family Spinosauridae, Baryonyx's affinities were obscure when it was discovered. Apart from the type species (B. walkeri), some researchers have suggested that Suchomimus tenerensis belongs in the same genus and that Suchosaurus cultridens is a senior synonym; subsequent authors have kept them separate. Baryonyx was the first theropod dinosaur demonstrated to have been piscivorous (fish-eating), as evidenced by fish scales in the stomach region of the holotype specimen. It may also have been an active predator of larger prey and a scavenger, since it also contained bones of a juvenile Iguanodon. The creature would have caught and processed its prey primarily with its forelimbs and large claws. Baryonyx lived near water bodies, in areas where other theropod, ornithopod, and sauropod dinosaurs have also been found.
Description

In 2010, Baryonyx was estimated to have been 7.5 m (25 ft) long and to have weighed 1.2 t (1.3 short tons).[1] It was estimated at 10 m (33 ft) in 1997, and 9.5 m (31 ft) long, 2.5 m (8.2 ft) in hip height, and 1.7 t (1.9 short tons) in weight in 1988. The fact that elements of the skull and vertebral column of the B. walkeri holotype specimen (NHM R9951) do not appear to have co-ossified (fused) suggests that the individual was not fully grown, and the mature animal may have been much larger (as attested by the size of the related Spinosaurus, which reached 15 m (49 ft) and 10 t (11 short tons). On the other hand, the specimen's fused sternum indicates that it may have been fairly mature.[2][3][4] The second-best-preserved specimen (ML1190) was about the same size as the holotype skeleton.[1][5]
The skull of Baryonyx is incompletely known, and much of the middle and hind portions are not preserved. The full length of the skull has been estimated to be 950 mm (37.4 in), based on comparison with that of the related genus Suchomimus (which is 20% larger). It was elongated, and the front 170 mm (6.6 in) of the premaxillae formed a long, low snout (rostrum) with a rounded upper surface. The nostrils, far back from the tip, passed horizontally from one side of the skull to the other. The front 130 mm (5.1 in) of the snout expanded into a spatulate (flared outwards to the sides), "terminal rosette" shape similar to the modern gharial, and the front 70 mm (2.7 in) of the lower margin was downturned. The snout was very narrow just behind the rosette. The creature's maxilla and premaxilla fit together in a complex articulation, resulting in a strongly curved tooth row. The gap in the row is comparable to that of Dilophosaurus. The front 140 mm (5.5 in) of the dentary in the mandible curved upwards towards this area, and the gap between the upper and lower jaw is known as the subrostral notch. The snout had extensive pits (which would have been exits for blood vessels and nerves), and the maxilla appears to have housed sinuses.[2][6]

Baryonyx had a rudimentary secondary palate, similar to crocodiles but unlike most theropod dinosaurs.[7] A rugose (roughly wrinkled) surface suggests the presence of a horny pad in the roof of the mouth. The upper midline of the nasal bones had a triangular sagittal crest, which was narrow and sharp in front. The lacrimal bone in front of the eye appears to have formed a horn core similar to those seen, for example, on Allosaurus. The dentary was very long and shallow, with a prominent Meckelian groove. The rest of the lower jaw was fragile; the hind third was much thinner than the front, with a blade-like appearance. The front part of the dentary curved outwards to accommodate the large front teeth, and this area formed the mandibular part of the rosette. The dentary had many foramina (openings), which were passages for nerves and blood vessels.[2] It has been suggested that some of Baryonyx's cranial bones had been misidentified (resulting in the occiput's too-deep reconstruction), and the skull was probably as low, long and narrow as that of the closely related Suchomimus.[8]

Most of the teeth found with the holotype specimen were not attached to the skull; a few remained in the upper jaw, and only small replacement teeth were in the lower jaw. The teeth had the shape of recurved cones, flattened somewhat sideways. The larger teeth were less recurved than the smaller ones, but were otherwise uniform. The roots were very long, and the teeth slender. The carinae (edges) of the teeth were finely serrated with denticles on the front and back. There were seven narrow, uniform denticles per millimetre (0.039 in), more than in most theropods. Some of the teeth were fluted, with six to eight ridges along the length of their inner sides and fine-grained enamel. The inner side of each tooth row had a bony wall. The number of teeth was large, with seven teeth in the right premaxilla (other theropods have three to five) and thirty-two in the dentary, where sixteen is typical. The lower jaw would have had sixty-four teeth, and the difference between the number of teeth in the upper and lower jaws is more pronounced than in other theropods. The teeth in the dentary were more densely packed than those in the maxilla, and probably smaller. The terminal rosette in the upper jaw had thirteen dental alveoli (tooth sockets), six on the left and seven on the right side; the first four were large (with the second and third the largest), while the fourth and fifth progressively decreased in size. The diameter of the largest was twice that of the smallest. The first four alveoli of the dentary (corresponding to the tip of the upper jaw) were the largest, with the rest more regular in size. Interdental plates were between the alveoli.[2][6]
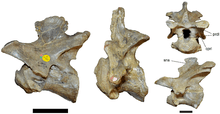
The neck formed a straighter S shape (a sigmoid curve typical of theropods) than that seen in other theropods; in fact, the neck was initially thought to lack the S curve.[9] The shape of the cervical vertebrae indicate that they tapered towards the head and were progressively longer front to back. The neural spines of the cervical vertebrae were low, thin, and were not always sutured to the centra (the bodies of the vertebrae). The axis vertebra, small relative to the size of the skull, had a well-developed hyposphene. The centra of the dorsal vertebrae were similar in size. Like other dinosaurs, Baryonyx reduced its weight (skeletal pneumaticity) with fenestrae (openings) in the neural arches and with pleurocoels (hollow depressions) in the centra (primarily near the transverse processes). From front to back, the neural spines of the dorsal vertebrae changed from short and stout to tall and broad.[2]
The scapulae (shoulder blades) were robust; the bones of the forelimb were short in relation to the animal's size, but broad and sturdy. The humerus was short and stout, with its ends broadly expanded and flattened—the upper side for the deltopectoral crest and muscle attachment and the lower for articulation with the radius and ulna. The radius was short, stout and straight, and the olecranon of the ulna apparently very powerful. The lower part of the ulna had a broad expansion. The first finger had a large claw (ungual bone) measuring about 31 cm (12 in) along its curve, which would have been lengthened by a keratin sheath in life. Apart from its size, the claw's proportions were fairly typical of a theropod; it was bilaterally symmetric, slightly compressed, smoothly rounded, and sharply pointed. A groove for the sheath ran along the length of the claw. The pubic foot of the pelvis was not expanded.[2]
History of discovery

On 7 January 1983 amateur fossil hunter William J. Walker discovered a large claw, a phalanx bone, and part of a rib in Smokejacks Pit, a clay pit near Ockley in Surrey, England. The tip of the claw was missing, but Walker found it a week later. British palaeontologists Alan J. Charig and Angela C. Milner examined the finds at the Natural History Museum of London and found more bones at the site on 7 February, but the entire skeleton could not be collected until May and June due to conditions at the pit. A team of eight museum staff members and several volunteers excavated two tonnes of matrix. Walker donated the claw to the museum, and the Ockley Brick Company (owners of the pit) donated the rest of the skeleton and provided equipment.[2][10] The area had been explored for 200 years, but no similar remains had been found before.[11]
Most of the bones collected were encased in siltstone nodules surrounded by fine sand and silt, with the rest lying in clay. The bones were disarticulated and scattered over a 5 x 2 m (17 x 8 ft) area, but most were not far from their natural positions. The position of some bones was disturbed by a bulldozer, and some were broken by mechanical equipment before they were collected. Preparing the specimen was difficult, due to the hardness of the siltstone matrix and the presence of siderite; acid preparation was attempted, but most of the matrix was removed mechanically. The skeleton consisted of partial skull bones; teeth; cervical, dorsal and caudal vertebrae; ribs; a sternum; coracoids; arm and hand bones; claws; hip bones, and leg bones.[2][12] The original specimen number was BMNH R9951, but it was later re-catalogued as NHMUK VP R9951.[12][13]

In 1986 Charig and Milner made the skeleton the holotype specimen of a new genus and species: Baryonyx walkeri. The genus name derives from ancient Greek; βαρύς (barys) means "heavy" or "strong", and ὄνυξ (onyx) means "claw" or "talon". The specific name honours Walker, for discovering the specimen. At that time, the authors did not know if the large claw belonged to the hand or the foot (as in dromaeosaurs, which it was then assumed to be). Due to ongoing work on the bones (70 percent had been prepared at the time), they called their article preliminary (a "Letter to Nature") and promised a more detailed description at a later date. Baryonyx was the first large Early Cretaceous theropod found anywhere in the world by that time.[12][14] Before the discovery of Baryonyx the last significant theropod find in the United Kingdom was Eustreptospondylus in 1871, and in a 1986 interview Charig called Baryonyx "the best find of the century" in Europe.[2][10] It was widely featured in international media, and its discovery was the subject of a 1987 BBC documentary. Baryonyx was nicknamed "Claws" by journalists punning on the title of the film Jaws. The skeleton is mounted at the Natural History Museum in London, and in 1997 Charig and Milner published a monograph describing the holotype skeleton in detail.[2][11]
Fossils from other parts of the UK and Iberia, mostly isolated teeth, have subsequently been attributed to Baryonyx or similar animals.[2] Isolated teeth and bones from the Isle of Wight, including hand bones and a vertebra, have been attributed to this genus.[15] A maxilla fragment from La Rioja, Spain, was attributed in 1995.[16] In 1999 a postorbital bone, a squamosal bone, a tooth, vertebra remains, metacarpals, and a phalanx from the Sala de los Infantes deposit in Burgos Province, Spain, were attributed to an immature Baryonyx (though some of these elements are unknown in the holotype),[17][18] and dinosaur tracks near Burgos have been identified as those of Baryonyx or a similar theropod.[19] In 2011 a specimen (ML1190) from the Papo Seco Formation in Boca do Chapim, Portugal, with a fragmentary dentary, teeth, vertebrae, ribs, hip bones, a scapula, and a phalanx bone, was attributed to Baryonyx, the most complete Iberian remains of the animal. The skeletal elements of this specimen are also represented in the more complete holotype NHM R9951, except for the mid-neck vertebrae.[5]
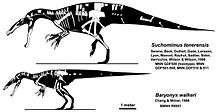
The authors of a 2002 article about the spinosaur Irritator proposed that Suchomimus tenerensis was similar enough to B. walkeri to be considered a species within the same genus (B. tenerensis), and suggested that Suchomimus was identical to Cristatusaurus; both are from the Elrhaz Formation of Niger.[20] At about 9.5 m (30 ft) and 2.5 tonnes (5,511 lb), Suchomimus was larger than Baryonyx.[1] In a 2004 conference abstract, palaeontologists Steve Hutt and Penny Newbery supported this view based on a large theropod vertebra from the Isle of Wight which they attributed to Baryonyx; this indicated that the vertebrae of the two genera were more similar than previously thought.[21] Later studies have kept the genera separate.[5][22][23]
In a 2003 article, Milner noted that the teeth of Baryonyx were very similar to those of the genus Suchosaurus and suggested that their remains represented the same animal.[24] The type species of the genus, S. cultridens, was named in 1841 based on teeth from Tilgate Forest in Sussex; a second species, S. girardi, was named in 1897 based on jaw fragments and a tooth from Boca do Chapim. In 2007 Buffetaut considered the teeth of S. girardi very similar to those of Baryonyx (and S. cultridens) except for the stronger development of the crown ribs, suggesting that the remains belonged to the same genus. Buffetaut agreed with Milner that the teeth of S. cultridens were almost identical to those of B. walkeri, but with a ribbier surface. The former taxon might be a senior synonym of the latter (since it was published first), depending on whether the differences were within a taxon or between different ones. According to Buffetaut, since the holotype specimen of S. cultridens is one worn tooth and that of B. walkeri is a skeleton it would be more practical to retain the newer name.[25] In 2011 Portuguese palaeontologist Octávio Mateus and colleagues agreed that Suchosaurus was closely related to Baryonyx, but considered both species in the former genus nomina dubia (dubious names) since their holotype specimens were not considered diagnostic (lacking distinguishing features) and could not be definitely equated with other taxa.[5]
Classification

In their original description, Charig and Milner found Baryonyx unique enough to warrant a new family of theropod dinosaurs: Baryonychidae. They found Baryonyx to be unlike any other theropod group (and considered the possibility that it was a thecodont, due to apparently primitive features), but noted that the articulation of the maxilla and premaxilla was similar to that in Dilophosaurus. They also noted that two fragmentary snouts from Niger, assigned to the family Spinosauridae by French palaeontologist Philippe Taquet in 1984, appeared almost identical to those of Baryonyx and they referred them to Baryonychidae instead.[12] In 1988, American palaeontologist Gregory S. Paul agreed with Taquet that Spinosaurus, described in 1915 based on fragmentary remains from Egypt which were destroyed in World War II, and Baryonyx were similar and (due to their kinked snouts) possibly late-surviving dilophosaurs.[3] French palaeontologist Eric Buffetaut also supported this relationship in 1989.[26] In 1990 Charig and Milner dismissed the spinosaurid affinities of Baryonyx, since they did not find their remains similar enough.[27]
Discoveries in the 1990s shed more light on the relationships of Baryonyx and its relatives. A snout from Morocco was referred to Spinosaurus and Irritator from Brazil was named in 1996.[28] Two years later the snout fragments from Niger were named Cristatusaurus, and Suchomimus was named from a partial skeleton from the country. In their description of Suchomimus, Sereno and colleagues placed it and Baryonyx in the new subfamily Baryonychinae within Spinosauridae; other members of the group were placed in the subfamily Spinosaurinae.[8][29] They also united the spinosaurids and their closest relatives in the superfamily Spinosauroidea, but in 2010 Roger Benson considered this a junior synonym of Megalosauroidea (an older name).[30] The following cladogram shows the position of Baryonyx within Spinosauridae, after Ronan Allain and colleagues, 2012:[22]

| Spinosauridae |
| ||||||||||||||||||||||||
| |
Evolution

Spinosaurids appear to have been widespread from the Barremian to the Cenomanian ages of the Cretaceous, about 130 to 95 million years ago. They shared features such as long, narrow, crocodile-like skulls; sub-circular teeth, with fine to no serrations; the snout rosette, and a secondary palate which made them more resistant to torsion. In contrast, the primitive and typical condition for theropods was a tall, narrow snout with blade-like teeth serrated front and back. The skull adaptations of spinosaurids converged with those of crocodilians; early members of the latter group had skulls similar to typical theropods, later developing elongated snouts, conical teeth, and secondary palates. These adaptations may have been the result of a dietary change from terrestrial prey to fish. Unlike crocodiles, the post-cranial skeletons of most spinosaurids (except Spinosaurus) do not appear to have aquatic adaptations.[31][4] Sereno and colleagues proposed that the large thumb-claw and robust forelimbs of spinosaurids evolved in the Middle Jurassic, before the elongation of the skull and other adaptations related to fish-eating, since the former features are shared with their megalosaurid relatives. They also suggested that the spinosaurines and baryonychines diverged before the Barremian age of the Early Cretaceous.[8]
Several theories have been proposed about the biogeography of the spinosaurids. Since Suchomimus was more closely related to Baryonyx (from Europe) than to Spinosaurus—although that genus also lived in Africa—the distribution of spinosaurids cannot be explained as vicariance resulting from continental rifting. Sereno et al. proposed that spinosaurids were initially distributed across the supercontinent Pangea, but split with the opening of the Tethys Sea. Spinosaurines would then have evolved in the south (Africa and South America: in Gondwanaland) and baryonychines in the north (Europe: in Laurasia), with Suchomimus the result of a single north-to-south dispersal event.[8] It has also been suggested that baryonychines could be the ancestors of spinosaurines, which appear to have replaced the former in Africa.[32] In 2006, it was demonstrated that Iberia was near northern Africa during the Early Cretaceous; some researchers have argued that the Iberian region was a stepping stone between Europe and Africa, which is supported by the presence of baryonychines in Iberia. The direction of the dispersal between Europe and Africa is still unknown,[25] and subsequent discoveries of spinosaurid remains in Asia and Australia indicate that it may have been complex.[5]
Palaeobiology

In 1986 Charig and Milner first suggested that its elongated snout with many finely serrated teeth indicated that Baryonyx was piscivorous (a fish-eater), speculating that it crouched on a riverbank and used its claw to gaff fish out of the water (similar to the modern grizzly bear). In 1984, Taquet pointed out that the spinosaurid snouts from Niger were similar to those of the modern gharial and suggested a behaviour similar to herons or storks. Charig and Milner did not consider Baryonyx to be aquatic (due to its nostrils being on the sides of its snout—far from the tip—and the form of the post-cranial skeleton), but thought it was capable of swimming like most land vertebrates. They later rejected their initial proposal that the articulation between the premaxilla and maxilla was mobile.[2][12]
In 1987 Andrew Kitchener disputed the piscivorous behaviour of Baryonyx and suggested that it would have been a scavenger, using its long neck to feed on the ground, its claws to break into a carcass, and its long snout (with nostrils far back for breathing) for investigating the body cavity. Kitchener argued that Baryonyx's jaws and teeth were too weak to kill other dinosaurs and too heavy to catch fish, with too many adaptations for piscivory.[33] According to R. E. H. Reid, a scavenged carcass would have been broken up by its predator and large animals capable of doing so—such as grizzly bears—are also capable of catching fish (at least in shallow water).[34]

In 1997, Charig and Milner demonstrated direct dietary evidence in the stomach region of the B. walkeri holotype. It contained the first evidence of piscivory in a theropod dinosaur, acid-etched scales and teeth of the common fish Scheenstia mantelli (then classified in the genus Lepidotes), and abraded bones of a young Iguanodon. An apparent gastrolith (gizzard stone) was also found. They also presented circumstantial evidence for piscivory, such as crocodile-like adaptations for catching and swallowing prey: long, narrow jaws with their "terminal rosette", similar to those of a gharial, and the downturned tip and notch of the snout. In their view, these adaptations suggested that Baryonyx would have caught small to medium-sized fish in the manner of a crocodilian: gripping them with the notch of the snout (giving the teeth a "stabbing function"), tilting the head backwards, and swallowing them headfirst. Larger fish would be broken up with the claws. That the teeth in the lower jaw were smaller, more crowded and numerous than those in the upper jaw may have helped the animal grip food. Charig and Milner maintained that Baryonyx would primarily have eaten fish (although it would also have been an active predator and opportunistic scavenger), but it was not equipped to be a macro-predator like Allosaurus. They suggested that Baryonyx mainly used its forelimbs and large claws to catch, kill and tear apart larger prey.[2][35] In 2004, a pterosaur neck vertebra from Brazil with a spinosaurid tooth embedded in it confirmed that the latter were not exclusively piscivorous.[36]

A 2007 finite element analysis of CT scanned snouts indicated that the biomechanics of Baryonyx were most similar to those of the gharial and unlike those of the American alligator and more-conventional theropods, supporting a piscivorous diet for spinosaurids.[7] Their secondary palate helped them resist bending and torsion of their tubular snouts.[7] A 2013 beam-theory study compared the biomechanics of CT-scanned spinosaurid snouts with those of extant crocodilians, and found the snouts of Baryonyx and Spinosaurus similar in their resistance to bending and torsion. Baryonyx was found to have relatively high resistance in the snout to dorsoventral bending compared with Spinosaurus and the gharial. The authors concluded (in contrast to the 2007 study) that Baryonyx performed differently than the gharial; spinosaurids were not exclusive piscivores, and their diet was determined by their individual size.[13]
A 2010 study proposed that spinosaurids were semi-aquatic, based on the oxygen isotope composition of spinosaurid teeth from around the world compared with that of other theropods and extant animals. Spinosaurids probably spent much of the day in water, like crocodiles and hippopotamuses, and had a diet similar to the former; both were opportunistic predators. Since most spinosaurids do not appear to have anatomical adaptations for an aquatic lifestyle, the authors proposed that submersion in water was a means of thermoregulation similar to that of crocodiles and hippopotamuses. Spinosaurids may also have turned to aquatic habitats and piscivory to avoid competition with large, more-terrestrial theropods.[37] A 2016 study found that the jaws of spinosaurids were convergent with those of pike conger eels; these fish also have sideways compressed jaws (whereas the jaws of crocodilians are compressed from top to bottom), an elongated snout with a "terminal rosette" that bears enlarged teeth, and a notch behind the rosette with smaller teeth. This type of jaws were likely evolved for grabbing prey in aquatic environments with low light, and may have helped in prey detection.[38]
In their original description Charig and Milner speculated that the elongated skull, long neck, and strong humerus of Baryonyx indicated that the animal was a facultative quadruped, unique among theropods.[12] In their 1997 article they found no skeletal support for this, but maintained thar the forelimbs would have been strong enough for a quadrupedal posture and it would probably have caught aquatic prey while crouching—or on all fours—near (or in) water.[2] A 2014 re-description of Spinosaurus based on new remains suggested that it was a quadruped, based on its anterior centre of body mass. The authors found quadrupedality unlikely for Baryonyx, since the better-known legs of the closely related Suchomimus did not support this posture.[4]
Palaeoecology

The Weald Clay formation consists of sediments of Hauterivian (Lower Weald Clay) to Barremian (Upper Weald Clay) in age, about 130–125 million years old. The B. walkeri holotype was found in the latter, in clay representing non-marine still water, which has been interpreted as a fluvial or mudflat environment with shallow water, lagoons, and marsh. During the Early Cretaceous, the Weald area of Surrey, Sussex, and Kent was partly covered by the large, fresh-to-brackish water Wealden Lake. Two large rivers drained the northern area (where London now stands), flowing into the lake through a river delta; the Anglo-Paris Basin was in the south. Its climate was sub-tropical, similar to the present Mediterranean region. Since the Smokejacks Pit consists of different stratigraphic levels, fossil taxa found there are not necessarily contemporaneous.[2] Dinosaurs from the locality include the ornithopods Mantellisaurus, Iguanodon, and small sauropods. Other vertebrates include sharks (such as Hybodus), bony fishes (including Scheenstia), crocodiles, and pterosaurs. Members of ten orders of insects have been identified, including Valditermes, Archisphex, and Pterinoblattina. Other invertebrates include ostracods, isopods, conchostracans, and bivalves. The plants Weichselia and the aquatic, herbaceous Bevhalstia were common. Other plants found include ferns, horsetails, club mosses, and conifers.[39]

Charig and Milner presented a possible scenario explaining the taphonomy (changes during fossilisation) of the B. walkeri holotype specimen. The fine-grained sediments around the skeleton, and the fact that the bones were found close together (skull and forelimb elements at one end of the excavation area and the pelvis and hind-limb elements at the other), indicates that the environment was quiet at the time of fossilisation and water currents did not carry the carcass far—possibly because the water was shallow. The area where the specimen died seems to have been suitable for a piscivorous animal. It may have caught fish and scavenged on the mud plain, becoming mired before it died and was buried. Since the bones are well-preserved and had no gnaw marks, the carcass appears to have been undisturbed by scavengers (suggesting that it was quickly covered by sediment). The disarticulation of the bones may have been the result of soft-tissue decomposition. Parts of the skeleton seem to have weathered to different degrees, perhaps because water levels changed or the sediments shifted (exposing parts of the skeleton). The girdle and limb bones, the dentary, and a rib were broken before fossilisation, perhaps from trampling by large animals while buried. The orientation of the bones indicates that the carcass lay on its back, which may explain why all the lower teeth had fallen out of their sockets and some upper teeth were still in place.[2]
Other dinosaurs from the Wessex Formation of the Isle of Wight include the theropods Neovenator, Aristosuchus, Thecocoelurus, Calamospondylus, and Ornithodesmus; the ornithopods Iguanodon, Hypsilophodon, and Valdosaurus; the sauropods Pelorosaurus and Chondrosteosaurus, and the ankylosaur Polacanthus.[40] The Papo Seco Formation of Portugal where Baryonyx has been identified is composed of marl, representing a lagoon environment. Other dinosaur remains from the area include fragments tentatively assigned to Mantellisaurus, a macronarian sauropod, and Megalosaurus. Most of the bones of Portuguese specimen ML1190 were damaged, and some scratches may be marks from small scavengers. The specimen's disarticulation indicates it was transported from a more-terrestrial environment (since many bones are missing), but those found were close together.[5][25]
See also
-
 Dinosaurs portal
Dinosaurs portal
References
- 1 2 3 Paul, G. S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press. pp. 87–88. ISBN 978-0-691-13720-9.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Charig, A. J.; Milner, A. C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum of London. 53: 11–70.
- 1 2 Paul, G. S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 271–274. ISBN 978-0-671-61946-6.
- 1 2 3 Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Lurino, D. A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–1616. Bibcode:2014Sci...345.1613I. doi:10.1126/science.1258750. PMID 25213375. Supplementary Information
- 1 2 3 4 5 6 Mateus, O.; Araújo, R.; Natário, C.; Castanhinha, R. (2011). "A new specimen of the theropod dinosaur Baryonyx from the early Cretaceous of Portugal and taxonomic validity of Suchosaurus" (PDF). Zootaxa. 2827: 54–68.
- 1 2 Hendrickx, C.; Mateus, O.; Buffetaut, E.; Evans, A. R. (2016). "Morphofunctional analysis of the quadrate of spinosauridae (Dinosauria: Theropoda) and the presence of Spinosaurus and a second spinosaurine taxon in the Cenomanian of North Africa". PLoS ONE. 11 (1): e0144695. Bibcode:2016PLoSO..1144695H. doi:10.1371/journal.pone.0144695. PMID 26734729.
- 1 2 3 Rayfield, E. J.; Milner, A. C.; Xuan, V. B.; Young, P. G. (2007). "Functional morphology of spinosaur 'crocodile-mimic' dinosaurs". Journal of Vertebrate Paleontology. 27 (4): 892–901. doi:10.1671/0272-4634(2007)27[892:FMOSCD]2.0.CO;2.
- 1 2 3 4 Sereno, P. C.; Beck, A. L.; Dutheil, D. B.; Gado, B.; Larsson, H. C. E.; Lyon, G. H.; Marcot, J. D.; Rauhut, O. W. M.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. D.; Wilson, G. P.; Wilson, J. A. (1998). "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids". Science. 282 (5392): 1298–1302. Bibcode:1998Sci...282.1298S. doi:10.1126/science.282.5392.1298. PMID 9812890. Retrieved 2013-03-19.
- ↑ Evers, S. W.; Rauhut, O. W. M.; Milner, A. C.; McFeeters, B.; Allain, R. (2015). "A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the "middle" Cretaceous of Morocco". PeerJ. 3: e1323. doi:10.7717/peerj.1323. PMC 4614847
 . PMID 26500829.
. PMID 26500829. - 1 2 Edwards, D. D. (1986). "Fossil Claw Unearths a New Family Tree". Science News. 130 (23): 356. doi:10.2307/3970849. JSTOR 3970849.
- 1 2 Moody, R. T. J.; Naish, D. (2010). "Alan Jack Charig (1927–1997): An overview of his academic accomplishments and role in the world of fossil reptile research". Geological Society, London, Special Publications. 343: 89–109. Bibcode:2010GSLSP.343...89M. doi:10.1144/SP343.6.
- 1 2 3 4 5 6 Charig, A. J.; Milner, A. C. (1986). "Baryonyx, a remarkable new theropod dinosaur". Nature. 324 (6095): 359–361. Bibcode:1986Natur.324..359C. doi:10.1038/324359a0. PMID 3785404.
- 1 2 Cuff, A. R.; Rayfield, E. J. (2013). Farke, Andrew A, ed. "Feeding Mechanics in Spinosaurid Theropods and Extant Crocodilians". PLoS ONE. 8 (5): e65295. Bibcode:2013PLoSO...865295C. doi:10.1371/journal.pone.0065295. PMC 3665537. PMID 23724135.
- ↑ Norman, D. B. (1985). "Dromaeosaurids". The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books. pp. 57–58. ISBN 978-0-517-46890-6.
- ↑ Clabby, S. M. (2005). "Baryonyx Charig and Milner 1986". DinoWight. Retrieved October 12, 2015.
- ↑ Viera, I.; Torres, J. A. (1995). "Presencia de Baryonyx walkeri (Saurischia, Theropoda) en el Weald de La Rioja (España)". Munibe Ciencias Naturales (in Spanish). 47: 57–61. ISSN 0214-7688.
- ↑ Vidarte, C. F.; Calvo, M. M.; Meijide, M.; Izquierdo, L. A.; Montero, D.; Pérez, G.; Torcida, F.; Urién, V.; Fuentes, F. M.; Fuentes, M. M. (2001). "Restos fósiles de Baryonyx (Dinosauria, Theropoda) en el Cretácico Inferior de Salas de los Infantes (Burgos, España)". Actas de las I Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos. (in Spanish): 349–359.
- ↑ Pereda-Suberbiola, X.; Ruiz-Omeñaca, J. I.; Canudo, J. I.; Torcida, F.; Sanz, J. L. (2012). "Dinosaur Faunas from the Early Cretaceous (Valanginian-Albian) of Spain". In Godefroit, P. Bernissart Dinosaurs. Indiana University Press. pp. 389–390. ISBN 978-0-253-00570-0.
- ↑ Pérez-Lorente, F. (2015). Dinosaur Footprints and Trackways of La Rioja. Life of the Past. Indiana: Indiana University Press. pp. 225–246. ISBN 978-0-253-01515-0.
- ↑ Sues, H. D.; Frey, E.; Martill, M.; Scott, D. M. (2002). "Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil". Journal of Vertebrate Paleontology. 22 (3): 535–547. doi:10.1671/0272-4634(2002)022[0535:icasdt]2.0.co;2.
- ↑ Hutt, S.; Newbery, P. (2004). "A new look at Baryonyx walkeri (Charig and Milner, 1986) based upon a recent fossil find from the Wealden". Symposium of Vertebrate Palaeontology and Comparative Anatomy.
- 1 2 Allain, R.; Xaisanavong, T.; Richir, P.; Khentavong, B. (2012). "The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos". Naturwissenschaften. 99 (5): 369–377. Bibcode:2012NW.....99..369A. doi:10.1007/s00114-012-0911-7. PMID 22528021.
- ↑ Benson, R. B. J.; Carrano, M. T.; Brusatte, S. L. (2009). "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic". Naturwissenschaften. 97 (1): 71–78. Bibcode:2010NW.....97...71B. doi:10.1007/s00114-009-0614-x. PMID 19826771.
- ↑ Milner, A. C. (2003). "Fish-eating theropods: A short review of the systematics, biology and palaeobiogeography of spinosaurs". Actas de las II Jornadas Internacionales sobre Paleontologýa de Dinosaurios y su Entorno: 129–138.
- 1 2 3 Buffetaut, E. (2007). "The spinosaurid dinosaur Baryonyx (Saurischia, Theropoda) in the Early Cretaceous of Portugal". Geological Magazine. 144 (6): 1021–1025. doi:10.1017/S0016756807003883.
- ↑ Buffetaut, E. (1989). "New remains of the enigmatic dinosaur Spinosaurus from the Cretaceous of Morocco and the affinities between Spinosaurus and Baryonyx". Neues Jahrbuch für Geologie und Paläontologie Monatshefte. 2: 79–87.
- ↑ Charig, A. J.; Milner, A. C. (1990). "The systematic position of Baryonyx walkeri, in the light of Gauthier's reclassification of the Theropoda". In Carpenter, K.; Currie, P. J. Dinosaur Systematics: Perspectives and Approaches. Cambridge: Cambridge University Press. pp. 127–140. ISBN 978-0-521-43810-0.
- ↑ Russell, D. A. (1996). "Isolated dinosaur bones from the Middle Cretaceous of the Tafilalt, Morocco". Bulletin du Muséum National d'Histoire Naturelle, Paris, 4e série, section C. 18 (2–3): 349–402.
- ↑ Taquet, P.; Russell, D. A. (1998). "New data on spinosaurid dinosaurs from the Early Cretaceous of the Sahara". Comptes Rendus de l'Académie des Sciences à Paris, Sciences de la Terre et des Planètes. 327 (5): 347–353. Bibcode:1998CRASE.327..347T. doi:10.1016/S1251-8050(98)80054-2.
- ↑ Benson, R. B. J. (2010). "A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods". Zoological Journal of the Linnean Society. 158 (4): 882–935. doi:10.1111/j.1096-3642.2009.00569.x.
- ↑ Holtz Jr., T. R. (1998). "Spinosaurs as crocodile mimics". Science. 282 (5392): 1276–1277. doi:10.1126/science.282.5392.1276.
- ↑ Buffetaut, E.; Ouaja, M. (2002). "A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae". Bulletin de la Société Géologique de France. 173 (5): 415–421. doi:10.2113/173.5.415.
- ↑ Kitchener, A. (1987). "Function of Claws' claws". Nature. 325 (6100): 114. Bibcode:1987Natur.325..114K. doi:10.1038/325114a0.
- ↑ Reid, R. E. H. (1987). "Claws' claws". Nature. 325 (6104): 487. Bibcode:1987Natur.325..487R. doi:10.1038/325487b0.
- ↑ López-Arbarello, A. (2012). "Phylogenetic Interrelationships of Ginglymodian Fishes (Actinopterygii: Neopterygii)". PLoS ONE. 7 (7): e39370. Bibcode:2012PLoSO...739370L. doi:10.1371/journal.pone.0039370. PMC 3394768. PMID 22808031.
- ↑ Buffetaut, E.; Martill, D.; Escuillié, F. (2004). "Pterosaurs as part of a spinosaur diet". Nature. 429 (6995): 33. Bibcode:2004Natur.429...33B. doi:10.1038/430033a. PMID 15229562.
- ↑ Amiot, R.; Buffetaut, E.; Lecuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, M. A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- ↑ Vullo, R.; Allain, R.; Cavin, L. (2016). "Convergent evolution of jaws between spinosaurid dinosaurs and pike conger eels". Acta Palaeontologica Polonica. 61. doi:10.4202/app.00284.2016.
- ↑ Ross, A. J.; Cook, E. (1995). "The stratigraphy and palaeontology of the Upper Weald Clay (Barremian) at Smokejacks Brickworks, Ockley, Surrey, England". Cretaceous Research. 16 (6): 705–716. doi:10.1006/cres.1995.1044.
- ↑ Martill, D. M.; Hutt, S. (1996). "Possible baryonychid dinosaur teeth from the Wessex Formation (Lower Cretaceous, Barremian) of the Isle of Wight, England". Proceedings of the Geologists' Association. 107 (2): 81–84. doi:10.1016/S0016-7878(96)80001-0.
External links
| Wikimedia Commons has media related to Baryonyx. |
| Wikispecies has information related to: Baryonyx |
- Natural History Museum – "Baryonyx: the discovery of an amazing fish-eating dinosaur" – four minute video presented by Angela Milner
| |||||||||||||||||||||||||||||||
| Piatnitzkysauridae |  | ||||||||||||||||||||||||||||||
| Megalosauria | |||||||||||||||||||||||||||||||