Tracheal cytotoxin

Tracheal cytotoxin (TCT) is a 921 dalton glycopeptide released by Bordetella pertussis and Neisseria gonorrhoeae.[1]
History
In 1980, it was discovered that B. pertussis could attach to hamster tracheal epithelial (HTE) cells, and also, that the supernatant from the cultured bacterium could disrupt the cell cycle of uninfected cells.[2] This prompted the scientists W. E. Goldman, D. G. Klapper, and J. B. Baseman to isolate and characterize a novel substance from B. pertussis supernatant. The novel disaccharide tetrapeptide that they had purified showed toxicity for HTE cells and tracheal ring cultures. Subsequently, they named the newly sequestered molecule tracheal cytotoxin (TCT).[3]
Structure

TCT is a soluble piece of peptidoglycan (PGN) found in the cell wall of all gram-negative bacteria.[4] Like all PGNs, TCT is composed of a disaccharide and a peptide chain. The IUPAC name for TCT is N-acetylglucosaminyl-1,6-anhydro-N-acetylmuramyl-(L)-alanyl-γ-(D)-glutamyl-mesodiaminopimelyl-(D)-alanine.[5] It is classified as a DAP(Diaminopimelic acid)-type PGN due to the third amino group within the chain being a diaminopimelyl peptide.

The DAP residue is responsible for directly bonding to the D-alanine peptide of another PGN molecule, thus aiding TCT's attachment within the cell wall.[6]
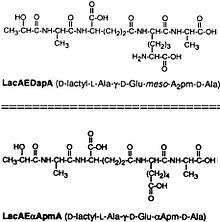
The DAP portion of TCT also implies importance in cytopathogenicity as analogs lacking DAP show a significant reduction in toxicity.[7]
Mechanism of Pathogenesis
Most gram-negative bacteria keep TCT within the cell wall by using a PGN-transporter protein known as AmpG. However, B. pertussis is not capable of recycling PGNs via AmpG and thus, TCT escapes into the surrounding environment.[8][9] Also, TCT appears to be constitutively expressed by B. pertussis.[4]
The first murine-model studies using TCT involved treatment of hamster tracheal cells. These experiments alluded to TCT's role in ciliostasis and cellular extrusion of ciliated hamster cells. Also, HTE cells had a markedly reduced level of DNA synthesis post-treatment with TCT.

While previous studies using murine models reported evidence of TCT causing ciliostasis, in vitro studies using human tracheal cells have shown that TCT does not affect ciliary beat frequency of living cells, but instead causes damage and eventual extrusion of ciliated cells.[10] In gonorrhea infections, vaginal ciliated epithelial cells have also displayed the same cytopathogenic effects due to TCT recognition.[11] The extensive damage to ciliated epithilial tissue caused by TCT results in major disruption to the ciliary escalator; an important asset of the host's non-specific defenses. This disruption hinders the host's ability to remove mucous and foreign microbes from the epithelial tissue. Paroxysmal cough, e.g. whooping cough, is a direct symptom of said mucous build-up due to ciliated tissue damage.
Effect on Immune System
TCT has been classified as an adjuvant molecule because of the stimulating effects it has on the immune system. Cellular damage associated with TCT is thought to be a result of increased levels of nitric oxide (NO) secretion by mucosal cells as part of an innate defense response to extracellular lipopolysaccharide (LPS) and TCT.[12] In humans, peptidoglycan recognition proteins, e.g. PGRPIαC, appear to bind with TCT and consequently induce the Tumor Necrosis Factor Receptor (TNFR) pathway.[13] Studies using murine macrophages have shown that TCT encourages cytokine secretion, probably through the Nod1 receptor.[14] As a pleiotropic toxin, TCT also acts as a pyrogen and as a stimulant of slow-wave sleep.[15]
References
- ↑ Cundell, DR; Kanthakumar, K; Taylor, GW; Goldman, WE; Flak, T; Cole, PJ; Wilson, R (1994). "Effect of tracheal cytotoxin from Bordetella pertussis on human neutrophil function in vitro". Infection and Immunity. 62 (2): 639–43. PMC 186151
 . PMID 8300220.
. PMID 8300220. - ↑ Goldman, W.E. and J.B. Baseman (March 1980). "Abstr. Ann. Meet. Am. Soc. Microbiol.". Abstr. Ann. Meet. Am. Soc. Microbiol.
- ↑ Goldman, William E.; Klapper, David G.; Baseman, Joel B. (1982-05-01). "Detection, isolation, and analysis of a released Bordetella pertussis product toxic to cultured tracheal cells". Infection and Immunity. 36 (2): 782–94. PMC 351298. PMID 6177637.
- 1 2 Mattoo, S.; Cherry, J. D. (2005). "Molecular Pathogenesis, Epidemiology, and Clinical Manifestations of Respiratory Infections Due to Bordetella pertussis and Other Bordetella Subspecies". Clinical Microbiology Reviews. 18 (2): 326–82. doi:10.1128/CMR.18.2.326-382.2005. PMC 1082800. PMID 15831828.
- ↑ Cookson, BT; Tyler, AN; Goldman, WE (1989). "Primary structure of the peptidoglycan-derived tracheal cytotoxin of Bordetella pertussis". Biochemistry. 28 (4): 1744–9. doi:10.1021/bi00430a048. PMID 2541765.
- ↑ Wilson, R; Read, R; Thomas, M; Rutman, A; Harrison, K; Lund, V; Cookson, B; Goldman, W; et al. (1991-01-01). "Effects of Bordetella pertussis infection on human respiratory epithelium in vivo and in vitro". Infection and Immunity. 59 (1): 337–45. PMC 257746. PMID 1987048.
- ↑ Luker, K. E.; Collier, J. L.; Kolodziej, E. W.; Marshall, G. R.; Goldman, W. E. (1993). "Bordetella pertussis Tracheal Cytotoxin and Other Muramyl Peptides: Distinct Structure-Activity Relationships for Respiratory Epithelial Cytopathology". Proceedings of the National Academy of Sciences. 90 (6): 2365–2369. doi:10.1073/pnas.90.6.2365.
- ↑ Jacobs, C; Joris, B; Jamin, M; Klarsov, K; Van Beeumen, J; Mengin-Lecreulx, D; Van Heijenoort, J; Park, JT; et al. (1995). "AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase". Molecular Microbiology. 15 (3): 553–9. doi:10.1111/j.1365-2958.1995.tb02268.x. PMID 7783625.
- ↑ Parkhill, Julian; Sebaihia, Mohammed; Preston, Andrew; Murphy, Lee D; Thomson, Nicholas; Harris, David E; Holden, Matthew T G; Churcher, Carol M; et al. (2003). "Comparative analysis of the genome sequences of Bordetella pertussis, Bordetella parapertussis and Bordetella bronchiseptica". Nature Genetics. 35 (1): 32–40. doi:10.1038/ng1227. PMID 12910271.
- ↑ Wilson, R; Read, R; Thomas, M; Rutman, A; Harrison, K; Lund, V; Cookson, B; Goldman, W; et al. (1991). "Effects of Bordetella pertussis infection on human respiratory epithelium in vivo and in vitro". Infection and immunity. 59 (1): 337–45. PMC 257746. PMID 1987048.
- ↑ Swaminathan, C. P.; Brown, P. H.; Roychowdhury, A.; Wang, Q.; Guan, R.; Silverman, N.; Goldman, W. E.; Boons, G.-J.; Mariuzza, R. A. (2006). "Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)". Proceedings of the National Academy of Sciences. 103 (3): 684–9. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
- ↑ Coote, JG (2001). "Environmental sensing mechanisms in Bordetella". Advances in microbial physiology. Advances in Microbial Physiology. 44: 141–81. doi:10.1016/S0065-2911(01)44013-6. ISBN 9780120277445. PMID 11407112.
- ↑ Swaminathan, C. P.; Brown, PH; Roychowdhury, A; Wang, Q; Guan, R; Silverman, N; Goldman, WE; Boons, GJ; Mariuzza, RA (2006). "Dual strategies for peptidoglycan discrimination by peptidoglycan recognition proteins (PGRPs)". Proceedings of the National Academy of Sciences. 103 (3): 684–9. doi:10.1073/pnas.0507656103. PMC 1334652. PMID 16407132.
- ↑ Humann, Jessica; Lenz, Laurel L. (2009). "Bacterial Peptidoglycan-Degrading Enzymes and Their Impact on Host Muropeptide Detection". Journal of Innate Immunity. 1 (2): 88–97. doi:10.1159/000181181. PMC 2659621. PMID 19319201.
- ↑ Magalhaes, Joao Gamelas; Philpott, Dana J; Nahori, Marie-Anne; Jéhanno, Muguette; Fritz, Joerg; Bourhis, Lionel Le; Viala, Jérôme; Hugot, Jean-Pierre; et al. (2005). "Murine Nod1 but not its human orthologue mediates innate immune detection of tracheal cytotoxin". EMBO Reports. 6 (12): 1201–7. doi:10.1038/sj.embor.7400552. PMC 1369207. PMID 16211083.