Chimpanzee genome project
The Chimpanzee Genome Project is an effort to determine the DNA sequence of the Chimpanzee genome. It is expected that by comparing the genomes of humans and other apes, it will be possible to better understand what makes humans distinct from other species from a genetic perspective. It will also aid in the study of diseases that affect (or, conversely, do not affect) various primate species.

Starting the chimpanzee genome project
Human and chimpanzee chromosomes are very similar. The primary difference is that humans have one fewer pair of chromosomes than do other great apes. Humans have 23 pairs of chromosomes and other great apes have 24 pairs of chromosomes. In the human evolutionary lineage, two ancestral ape chromosomes fused at their telomeres, producing human chromosome 2.[3] There are nine other major chromosomal differences between chimpanzees and humans: chromosome segment inversions on human chromosomes 1, 4, 5, 9, 12, 15, 16, 17, and 18. After the completion of the Human genome project, a common chimpanzee genome project was initiated. In December 2003, a preliminary analysis of 7600 genes shared between the two genomes confirmed that certain genes such as the forkhead-box P2 transcription factor, which is involved in speech development, are different in the human lineage. Several genes involved in hearing were also found to have changed during human evolution, suggesting selection involving human language-related behavior. Differences between individual humans and common chimpanzees are estimated to be about 10 times the typical difference between pairs of humans.[4]
Draft genome sequence of the common chimpanzee
Analysis of the genome was published in Nature on September 1, 2005, in an article produced by the Chimpanzee Sequencing and Analysis Consortium, a group of scientists which is supported in part by the National Human Genome Research Institute, one of the National Institutes of Health. The article marked the completion of the draft genome sequence.[4] A database [5] now exists containing the genetic differences between human and chimpanzee genes, with about thirty-five million single-nucleotide changes, five million insertion/deletion events, and various chromosomal rearrangements. Gene duplications account for most of the sequence differences between humans and chimps. Single-base-pair substitutions account for about half as much genetic change as does gene duplication.
Typical human and chimp homologs of proteins differ in only an average of two amino acids. About 30 percent of all human proteins are identical in sequence to the corresponding chimp protein. As mentioned above, gene duplications are a major source of differences between human and chimp genetic material, with about 2.7 percent of the genome now representing differences having been produced by gene duplications or deletions during approximately 6 million years [6] since humans and chimps diverged from their common evolutionary ancestor. The comparable variation within human populations is 0.5 percent.[7]
About 600 genes have been identified that may have been undergoing strong positive selection in the human and chimp lineages; many of these genes are involved in immune system defense against microbial disease (example: granulysin is protective against Mycobacterium tuberculosis [8]) or are targeted receptors of pathogenic microorganisms (example: Glycophorin C and Plasmodium falciparum). By comparing human and chimp genes to the genes of other mammals, it has been found that genes coding for transcription factors, such as forkhead-box P2 (FOXP2), have often evolved faster in the human relative to chimp; relatively small changes in these genes may account for the morphological differences between humans and chimps. A set of 348 transcription factor genes code for proteins with an average of about 50 percent more amino acid changes in the human lineage than in the chimp lineage.
Six human chromosomal regions were found that may have been under particularly strong and coordinated selection during the past 250,000 years. These regions contain at least one marker allele that seems unique to the human lineage while the entire chromosomal region shows lower than normal genetic variation. This pattern suggests that one or a few strongly selected genes in the chromosome region may have been preventing the random accumulation of neutral changes in other nearby genes. One such region on chromosome 7 contains the FOXP2 gene (mentioned above) and this region also includes the Cystic fibrosis transmembrane conductance regulator (CFTR) gene, which is important for ion transport in tissues such as the salt-secreting epithelium of sweat glands. Human mutations in the CFTR gene might be selected for as a way to survive cholera.[9]
Another such region on chromosome 4 may contain elements regulating the expression of a nearby protocadherin gene that may be important for brain development and function. Although changes in expression of genes that are expressed in the brain tend to be less than for other organs (such as liver) on average, gene expression changes in the brain have been more dramatic in the human lineage than in the chimp lineage.[10] This is consistent with the dramatic divergence of the unique pattern of human brain development seen in the human lineage compared to the ancestral great ape pattern. The protocadherin-beta gene cluster on chromosome 5 also shows evidence of possible positive selection.[11]
Results from the human and chimp genome analyses should help in understanding some human diseases. Humans appear to have lost a functional caspase-12 gene, which in other primates codes for an enzyme that may protect against Alzheimer's disease.
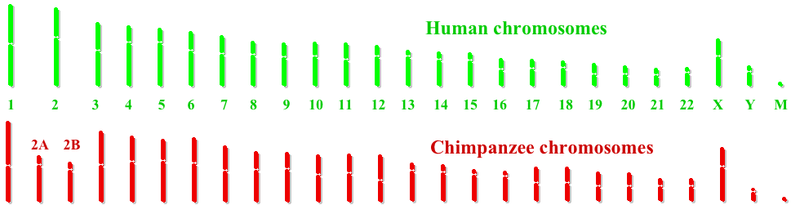
Genes of the chromosome 2 fusion site

The results of the chimpanzee genome project suggest that when ancestral chromosomes 2A and 2B fused to produce human chromosome 2, no genes were lost from the fused ends of 2A and 2B. At the site of fusion, there are approximately 150,000 base pairs of sequence not found in chimpanzee chromosomes 2A and 2B. Additional linked copies of the PGML/FOXD/CBWD genes exist elsewhere in the human genome, particularly near the p end of chromosome 9. This suggests that a copy of these genes may have been added to the end of the ancestral 2A or 2B prior to the fusion event. It remains to be determined if these inserted genes confer a selective advantage.
- PGML. The phosphoglucomutase-like gene of human chromosome 2. This gene is incomplete and may not produce a functional transcript.[12]
- FOXD. The forkhead box D4-like gene is an example of an intronless gene. The function of this gene is not known, but it may code for a transcription control protein.
- CBWD. Cobalamin synthetase is a bacterial enzyme that makes vitamin B12. In the distant past, a common ancestor to mice and apes incorporated a copy of a cobalamin synthetase gene (see: Horizontal gene transfer). Humans are unusual in that they have several copies of cobalamin synthetase-like genes, including the one on chromosome 2. It remains to be determined what the function of these human cobalamin synthetase-like genes is. If these genes are involved in vitamin B12 metabolism, this could be relevant to human evolution. A major change in human development is greater post-natal brain growth than is observed in other apes. Vitamin B12 is important for brain development, and vitamin B12 deficiency during brain development results in severe neurological defects in human children.
- CXYorf1-like protein. Several transcripts of unknown function corresponding to this region have been isolated. This region is also present in the closely related chromosome 9p terminal region that contains copies of the PGML/FOXD/CBWD genes.
- Many ribosomal protein L23a pseudogenes are scattered through the human genome.
See also
| Wikiversity has learning materials about Chimpanzee Genome Project |
References
- ↑ McConkey EH (2004). "Orthologous numbering of great ape and human chromosomes is essential for comparative genomics". Cytogenet. Genome Res. 105 (1): 157–8. doi:10.1159/000078022. PMID 15218271.
- ↑ Springer MS, Murphy WJ, Eizirik E, O'Brien SJ; Murphy; Eizirik; O'Brien (February 2003). "Placental mammal diversification and the Cretaceous-Tertiary boundary". Proc. Natl. Acad. Sci. U.S.A. 100 (3): 1056–61. Bibcode:2003PNAS..100.1056S. doi:10.1073/pnas.0334222100. PMC 298725
 . PMID 12552136.
. PMID 12552136. - ↑ De Grouchy J (August 1987). "Chromosome phylogenies of man, great apes, and Old World monkeys". Genetica. 73 (1–2): 37–52. doi:10.1007/bf00057436. PMID 3333352.
- 1 2 Chimpanzee Sequencing; Analysis Consortium (2005). "Initial sequence of the chimpanzee genome and comparison with the human genome" (PDF). Nature. 437 (7055): 69–87. Bibcode:2005Natur.437...69.. doi:10.1038/nature04072. PMID 16136131.
- ↑ "Chimpanzee genome database".
- ↑ Caswell JL, Mallick S, Richter DJ, Neubauer J, Schirmer C, Gnerre S, Reich D (2008-04-18). McVean, Gil, ed. "Analysis of Chimpanzee History Based on Genome Sequence Alignments". PLoS Genet. 4 (4): e1000057. doi:10.1371/journal.pgen.1000057. PMC 2278377. PMID 18421364.
- ↑ Cheng Z, Ventura M, et al. (2005). "A genome-wide comparison of recent chimpanzee and human segmental duplications". Nature. 437 (7055): 88–93. Bibcode:2005Natur.437...88C. doi:10.1038/nature04000. PMID 16136132.
- ↑ Stenger S, Hanson DA, Teitelbaum R; Hanson; Teitelbaum; Dewan; Niazi; Froelich; Ganz; Thoma-Uszynski; Melian; Bogdan; Porcelli; Bloom; Krensky; Modlin; et al. (October 1998). "An antimicrobial activity of cytolytic T cells mediated by granulysin". Science. 282 (5386): 121–5. Bibcode:1998Sci...282..121S. doi:10.1126/science.282.5386.121. PMID 9756476.
- ↑ Goodman BE, Percy WH (June 2005). "CFTR in cystic fibrosis and cholera: from membrane transport to clinical practice". Adv Physiol Educ. 29 (2): 75–82. doi:10.1152/advan.00035.2004. PMID 15905150.
- ↑ Khaitovich; Hellmann, I; Enard, W; Nowick, K; Leinweber, M; Franz, H; Weiss, G; Lachmann, M; Pääbo, S; et al. (2005). "Parallel Patterns of Evolution in the Genomes and Transcriptomes of Humans and Chimpanzees". Science. 309 (5742): 1850–4. Bibcode:2005Sci...309.1850K. doi:10.1126/science.1108296. PMID 16141373.
- ↑ Miki R, Hattori K, Taguchi Y, et al. (April 2005). "Identification and characterization of coding single-nucleotide polymorphisms within human protocadherin-alpha and -beta gene clusters". Gene. 349: 1–14. doi:10.1016/j.gene.2004.11.044. PMID 15777644.
- ↑ Fan Y, Newman T, Linardopoulou E, Trask BJ (November 2002). "Gene content and function of the ancestral chromosome fusion site in human chromosome 2q13-2q14.1 and paralogous regions". Genome Res. 12 (11): 1663–72. doi:10.1101/gr.338402. PMC 187549. PMID 12421752.
| Ape species |
| |
|---|---|---|
| Ape study | ||
| Legal and social status | ||
| See also |
| |
