Wnt signaling pathway
The Wnt signaling pathways are a group of signal transduction pathways made of proteins that pass signals into a cell through cell surface receptors. Three Wnt signaling pathways have been characterized: the canonical Wnt pathway, the noncanonical planar cell polarity pathway, and the noncanonical Wnt/calcium pathway. All three pathways are activated by binding a Wnt-protein ligand to a Frizzled family receptor, which passes the biological signal to the Dishevelled protein inside the cell. The canonical Wnt pathway leads to regulation of gene transcription. The noncanonical planar cell polarity pathway regulates the cytoskeleton that is responsible for the shape of the cell. The noncanonical Wnt/calcium pathway regulates calcium inside the cell. Wnt signaling pathways use either nearby cell-cell communication (paracrine) or same-cell communication (autocrine). They are highly evolutionarily conserved in animals, which means they are similar across animal species from fruit flies to humans.[1][2]
Wnt signaling was first identified for its role in carcinogenesis, then for its function in embryonic development. The embryonic processes it controls include body axis patterning, cell fate specification, cell proliferation and cell migration. These processes are necessary for proper formation of important tissues including bone, heart and muscle. Its role in embryonic development was discovered when genetic mutations in Wnt pathway proteins produced abnormal fruit fly embryos. Wnt signaling also controls tissue regeneration in adult bone marrow, skin and intestine.[3] Later research found that the genes responsible for these abnormalities also influenced breast cancer development in mice.
This pathway's clinical importance was demonstrated by mutations that lead to various diseases, including breast and prostate cancer, glioblastoma, type II diabetes and others.[4][5]
History
The discovery of Wnt signaling was influenced by research on oncogenic (cancer-causing) retroviruses. In 1982, Roel Nusse and Harold Varmus infected mice with mouse mammary tumor virus in order to mutate mouse genes to see which mutated genes could cause breast tumors. They identified a new mouse proto-oncogene that they named int1 (integration 1).[2][6]
Int1 is highly conserved across multiple species, including humans and Drosophila. Its presence in D. melanogaster led researchers to discover in 1987 that the int1 gene in Drosophila was actually the already known and characterized Drosophila gene known as Wingless (Wg).[2] Since previous research by Christiane Nüsslein-Volhard and Eric Wieschaus (which won them the Nobel Prize in Physiology or Medicine in 1995) had already established the function of Wg as a segment polarity gene involved in the formation of the body axis during embryonic development, researchers determined that the mammalian int1 discovered in mice is also involved in embryonic development.[7]
Etymology
Continued research led to the discovery of further int1-related genes; however, because those genes were not identified in the same manner as int1, the int gene nomenclature was inadequate. Thus, the int/Wingless family became the Wnt family and int1 became Wnt1. The name Wnt is a portmanteau of int and Wg and stands for "Wingless-related integration site".[2]
Proteins
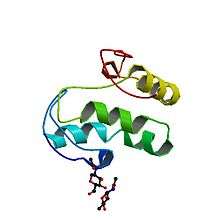
Wnt comprises a diverse family of secreted lipid-modified signaling glycoproteins that are 350–400 amino acids in length.[8] The type of lipid modification that occurs on these proteins is palmitoylation of cysteines in a conserved pattern of 23–24 cysteine residues.[4] Palmitoylation is necessary because it initiates targeting of the Wnt protein to the plasma membrane for secretion and it allows the Wnt protein to bind its receptor due to the covalent attachment of fatty acids. Wnt proteins also undergo glycosylation, which attaches a carbohydrate in order to ensure proper secretion.[9] In Wnt signaling, these proteins act as ligands to activate the different Wnt pathways via paracrine and autocrine routes.[1][5]
These proteins are highly conserved across species.[2] They can be found in mice, humans, Xenopus, zebrafish, Drosophila and many others.[10]
| Species | Wnt proteins |
|---|---|
| Homo sapiens | WNT1, WNT2, WNT2B, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16 |
| Mus musculus | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt9A, Wnt9B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Xenopus | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt11R |
| Danio rerio | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Drosophila | Wg, DWnt2, DWnt3/5, DWnt 4, DWnt6, WntD/DWnt8, DWnt10 |
| Hydra | hywnt1, hywnt5a, hywnt8, hywnt7, hywnt9/10a, hywnt9/10b, hywnt9/10c, hywnt11, hywnt16 |
Mechanism
Foundation
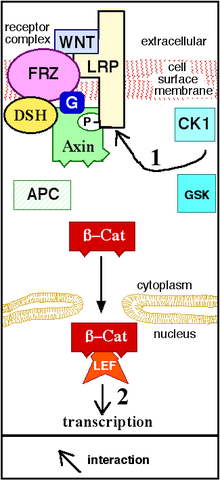
Wnt signaling begins when a Wnt protein binds to the N-terminal extra-cellular cysteine-rich domain of a Frizzled (Fz) family receptor.[11] These receptors span the plasma membrane seven times and constitute a distinct family of G-protein coupled receptors (GPCRs).[12] However, to facilitate Wnt signaling, co-receptors may be required alongside the interaction between the Wnt protein and Fz receptor. Examples include lipoprotein receptor-related protein (LRP)-5/6, receptor tyrosine kinase (RTK), and ROR2.[5] Upon activation of the receptor, a signal is sent to the phosphoprotein Dishevelled (Dsh), which is located in the cytoplasm. This signal is transmitted via a direct interaction between Fz and Dsh. Dsh proteins are present in all organisms and they all share the following highly conserved protein domains: an amino-terminal DIX domain, a central PDZ domain, and a carboxy-terminal DEP domain. These different domains are important because after Dsh, the Wnt signal can branch off into multiple pathways and each pathway interacts with a different combination of the three domains.[13]
Canonical and noncanonical pathways
The three best characterized Wnt signaling pathways are the canonical Wnt pathway, the noncanonical planar cell polarity pathway, and the noncanonical Wnt/calcium pathway. As their names suggest, these pathways belong to one of two categories: canonical or noncanonical. The difference between the categories is that a canonical pathway involves the protein β-catenin while a noncanonical pathway operates independently of it.[11]
.jpg)
Canonical pathway
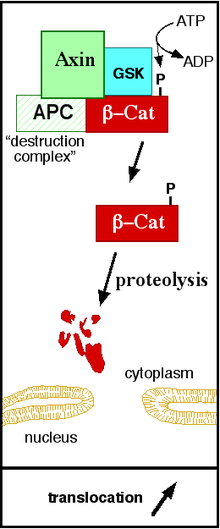
The canonical Wnt pathway (or Wnt/β-catenin pathway) is the Wnt pathway that causes an accumulation of β-catenin in the cytoplasm and its eventual translocation into the nucleus to act as a transcriptional coactivator of transcription factors that belong to the TCF/LEF family. Without Wnt signaling, β-catenin would not accumulate in the cytoplasm since a destruction complex would normally degrade it. This destruction complex includes the following proteins: Axin, adenomatosis polyposis coli (APC), protein phosphatase 2A (PP2A), glycogen synthase kinase 3 (GSK3) and casein kinase 1α (CK1α).[14][15] It degrades β-catenin by targeting it for ubiquitination, which subsequently sends it to the proteasome to be digested.[11][16] However, as soon as Wnt binds Fz and LRP5/6, the destruction complex function becomes disrupted. This is due to Wnt causing the translocation of the negative Wnt regulator, Axin, and the destruction complex to the plasma membrane. Phosphorylation by other proteins in the destruction complex subsequently binds Axin to the cytoplasmic tail of LRP5/6. Axin becomes de-phosphorylated and its stability and levels decrease. Dsh then becomes activated via phosphorylation and its DIX and PDZ domains inhibit the GSK3 activity of the destruction complex. This allows β-catenin to accumulate and localize to the nucleus and subsequently induce a cellular response via gene transduction alongside the TCF/LEF (T-cell factor/lymphoid enhancing factor)[17] transcription factors.[16]
Noncanonical pathways
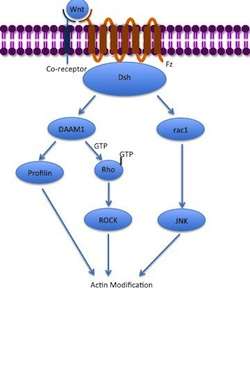
The noncanonical planar cell polarity (PCP) pathway does not involve β-catenin. It does not use LRP-5/6 as its co-receptor and is thought to use NRH1, Ryk, PTK7 or ROR2. The PCP pathway is activated via the binding of Wnt to Fz and its co-receptor. The receptor then recruits Dsh, which uses its PDZ and DIX domains to form a complex with Dishevelled-associated activator of morphogenesis 1 (DAAM1). Daam1 then activates the small G-protein Rho through a guanine exchange factor. Rho activates Rho-associated kinase (ROCK), which is one of the major regulators of the cytoskeleton. Dsh also forms a complex with rac1 and mediates profilin binding to actin. Rac1 activates JNK and can also lead to actin polymerization. Profilin binding to actin can result in restructuring of the cytoskeleton and gastrulation.[5][18]
.jpg)
The noncanonical Wnt/calcium pathway also does not involve β-catenin. Its role is to help regulate calcium release from the endoplasmic reticulum (ER) in order to control intracellular calcium levels. Like other Wnt pathways, upon ligand binding, the activated Fz receptor directly interacts with Dsh and activates specific Dsh-protein domains. The domains involved in Wnt/calcium signaling are the PDZ and DEP domains.[5] However, unlike other Wnt pathways, the Fz receptor directly interfaces with a trimeric G-protein. This co-stimulation of Dsh and the G-protein can lead to the activation of either PLC or cGMP-specific PDE. If PLC is activated, the plasma membrane component PIP2 is cleaved into DAG and IP3. When IP3 binds its receptor on the ER, calcium is released. Increased concentrations of calcium and DAG can activate Cdc42 through PKC. Cdc42 is an important regulator of ventral patterning. Increased calcium also activates calcineurin and CaMKII. CaMKII induces activation of the transcription factor NFAT, which regulates cell adhesion, migration and tissue separation.[5] Calcineurin activates TAK1 and NLK kinase, which can interfere with TCF/ß-Catenin signaling in the canonical Wnt pathway.[19] However, if PDE is activated, calcium release from the ER is inhibited. PDE mediates this through the inhibition of PKG, which subsequently causes the inhibition of calcium release.[5]
Integrated Wnt pathway
The binary distinction of canonical and non-canonical Wnt signaling pathways has come under scrutiny and an integrated, convergent Wnt pathway has been proposed.[20] Some evidence for this was found for one Wnt ligand (Wnt5A).[21] Evidence for a convergent Wnt signaling pathway that shows integrated activation of Wnt/Ca2+ and Wnt/ß-catenin signaling, for multiple Wnt ligands, was described in mammalian cell lines.[22]
Other pathways
Wnt signaling also regulates a number of other signaling pathways that have not been as extensively elucidated. One such pathway includes the interaction between Wnt and GSK3. During cell growth, Wnt can inhibit GSK3 in order to activate mTOR in the absence of β-catenin. However, Wnt can also serve as a negative regulator of mTOR via activation of the tumor suppressor TSC2, which is upregulated via Dsh and GSK3 interaction.[23] During myogenesis, Wnt uses PA and CREB to activate MyoD and Myf5 genes.[24] Wnt also acts in conjunction with Ryk and Src to allow for regulation of neuron repulsion during axonal guidance. Wnt regulates gastrulation when CK1 serves as an inhibitor of Rap1-ATPase in order to modulate the cytoskeleton during gastrulation. Further regulation of gastrulation is achieved when Wnt uses ROR2 along with the CDC42 and JNK pathway to regulate the expression of PAPC. Dsh can also interact with aPKC, Pa3, Par6 and LGl in order to control cell polarity and microtubule cytoskeleton development. While these pathways overlap with components associated with PCP and Wnt/Calcium signaling, they are considered distinct pathways because they produce different responses.[5]
Regulation
In order to ensure proper functioning, Wnt signaling is constantly regulated at several points along its signaling pathways.[25] For example, Wnt proteins are palmitoylated. The protein porcupine mediates this process, which means that it helps regulate when the Wnt ligand is secreted by determining when it is fully formed. Secretion is further controlled with proteins such as wntless and evenness interrupted and complexes such as the retromer complex.[5][16] Upon secretion, the ligand can be prevented from reaching its receptor through the binding of proteins such as the stabilizers Dally and glypican 3, which inhibit diffusion. At the Fz receptor, the binding of proteins other than Wnt can antagonize signaling. Specific antagonists include Dickkopf (Dkk), Wnt inhibitory factor 1 (WIF-1), secreted Frizzled-related proteins (SFRP), Cerberus, Frzb, Wise, and SOST. These constitute inhibitors of Wnt signaling. However, other molecules also act as activators. Norrin and R-Spondin2 activate Wnt signaling in the absence of Wnt ligand. Interactions between Wnt signaling pathways also regulate Wnt signling. As previously mentioned, the Wnt/calcium pathway can inhibit TCF/β-catenin, preventing canonical Wnt pathway signaling.[5][16] ProstaglandinE2 is an essential activator of the canonical Wnt signaling pathway. Interaction of PGE2 with its receptors E2/E4 stabilizes beta catenin through cAMP/PKA mediated phosphorylation. The synthesis of PGE2 is be necessary for Wnt signaling mediated processes such as tissue regeneration and control of stem cell population in zebrafish and mouse.[3] Intriguingly, the unstructured regions of several oversized Intrinsically disordered proteinsplay crucial roles in regulating Wnt signaling.[26]
Induced cell responses
Embryonic development
Wnt signaling plays a critical role in embryonic development. It operates in both vertebrates and invertebrates, including humans, frogs, zebrafish, C. elegans, Drosophila and others. It was first found in the segment polarity of Drosophila, where it helps to establish anterior and posterior polarities. It is implicated in other developmental processes. As its function in Drosophila suggests, it plays a key role in body axis formation, particularly the formation of the anteroposterior and dorsoventral axes. It is involved in the induction of cell differentiation to prompt formation of important organs such as lungs and ovaries. Wnt further ensures the development of these tissues through proper regulation of cell proliferation and migration. Wnt signaling functions can be divided into axis patterning, cell fate specification, cell proliferation and cell migration.[27]
Axis patterning
In early embryo development, the formation of the primary body axes is a crucial step in establishing the organism's overall body plan. The axes include the anteroposterior axis, dorsoventral axis, and right-left axis. Wnt signaling is implicated in the formation of the anteroposterior and dorsoventral (DV) axes. Wnt signaling activity in anterior-posterior development can be seen in mammals, fish and frogs. In mammals, the primitive streak and other surrounding tissues produce the morphogenic compounds Wnts, BMPs, FGFs, Nodal and retinoic acid to establish the posterior region during late gastrula. These proteins form concentration gradients. Areas of highest concentration establish the posterior region while areas of lowest concentration indicate the anterior region. In fish and frogs, β-catenin produced by canonical Wnt signaling causes the formation of organizing centers, which, alongside BMPs, elicit posterior formation. Wnt involvement in DV axis formation can be seen in the activity of the formation of the Spemann organizer, which establishes the dorsal region. Canonical Wnt signaling β-catenin production induces the formation of this organizer via the activation of the genes twin and siamois.[20][27] Similarly, in avian gastrulation, cells of the Koller's sickle express different mesodermal marker genes that allow for the differential movement of cells during the formation of the primitive streak. Wnt signaling activated by FGFs is responsible for this movement.[28][29]
Wnt signaling is also involved in the axis formation of specific body parts and organ systems later in development. In vertebrates, sonic hedgehog (Shh) and Wnt morphogenetic signaling gradients establish the dorsoventral axis of the central nervous system during neural tube axial patterning. High Wnt signaling establishes the dorsal region while high Shh signaling indicates the ventral region.[30] Wnt is involved in the DV formation of the central nervous system through its involvement in axon guidance. Wnt proteins guide the axons of the spinal cord in an anterior-posterior direction.[31] Wnt is also involved in the formation of the limb DV axis. Specifically, Wnt7a helps produce the dorsal patterning of the developing limb.[20][27]
Cell fate specification
Cell fate specification or cell differentiation is a process where undifferentiated cells can become a more specialized cell type. Wnt signaling induces differentiation of pluripotent stem cells into mesoderm and endoderm progenitor cells.[32] These progenitor cells further differentiate into cell types such as endothelial, cardiac and vascular smooth muscle lineages.[33] Wnt signaling induces blood formation from stem cells. Specifically, Wnt3 leads to mesoderm committed cells with hematopoietic potential.[34] Wnt1 antagonizes neural differentiation and is a major factor in self-renewal of neural stem cells. This allows for regeneration of nervous system cells, which is further evidence of a role in promoting neural stem cell proliferation.[32] Wnt signaling is involved in germ cell determination, gut tissue specification, hair follicle development, lung tissue development, trunk neural crest cell differentiation, nephron development, ovary development and sex determination.[27]
Cell proliferation
In order to have the mass differentiation of cells needed to form the specified cell tissues of different organisms, proliferation and growth of embryonic stem cells must take place. This process is mediated through canonical Wnt signaling, which increases nuclear and cytoplasmic β-catenin. Increased β-catenin can initiate transcriptional activation of proteins such as cyclin D1 and c-myc, which control the G1 to S phase transition in the cell cycle. Entry into the S phase causes DNA replication and ultimately mitosis, which are responsible for cell proliferation.[35] This proliferation increase is directly paired with cell differentiation because as the stem cells proliferate, they also differentiate. This allows for overall growth and development of specific tissue systems during embryonic development. This is apparent in systems such as the circulatory system where Wnt3a leads to proliferation and expansion of hematopoietic stem cells needed for red blood cell formation.[36]
The biochemistry of cancer stem cells is subtly different than that of other tumor cells. These so-called Wnt-addicted cells hijack and depend on constant stimulation of the Wnt pathway to promote their uncontrolled growth, survival and migration. In cancer, Wnt signaling can become independent of regular stimuli, through mutations in downstream oncogenes and tumor suppressor genes that become permanently activated even though the normal receptor has not received a signal. β-catenin binds to transcription factors such as the protein TCF4 and in combination the molecules activate the necessary genes. LF3 strongly inhibits this binding in vitro, in cell lines and reduced tumor growth in mouse models. It prevented replication and reduced their ability to migrate, all without affecting healthy cells. No cancer stem cells remained after treatment. The discovery was the product of "rational drug design", involving AlphaScreens and ELISA technologies.[37]
Cell migration
Cell migration during embryonic development allows for the establishment of body axes, tissue formation, limb induction and several other processes. Wnt signaling helps mediate this process, particularly during convergent extension. Signaling from both the Wnt PCP pathway and canonical Wnt pathway is required for proper convergent extension during gastrulation. Convergent extension is further regulated by the Wnt/calcium pathway, which blocks convergent extension when activated. Wnt signaling also induces cell migration in later stages of development through the control of the migration behavior of neuroblasts, neural crest cells, myocytes, and tracheal cells.[38]
Wnt signaling is involved in another key migration process known as the epithelial-mesenchymal transition (EMT). This process allows epithelial cells to transform into mesenchymal cells so that they are no longer held in place at the laminin. It involves cadherin down-regulation so that cells can detach from laminin and migrate. Wnt signaling is an inducer of EMT, particularly in mammary development.[39]
Insulin sensitivity
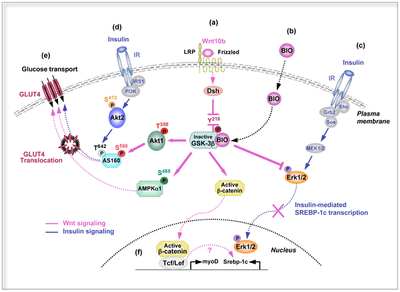
Insulin is a peptide hormone involved in glucose homeostasis within certain organisms. Specifically, it leads to upregulation of glucose transporters in the cell membrane in order to increase glucose uptake from the bloodstream. This process is partially mediated by activation of Wnt/β-catenin signaling, which can increase a cell's insulin sensitivity. In particular, Wnt10b is a Wnt protein that increases this sensitivity in skeletal muscle cells.[40]
Clinical implications
Cancer
Since its initial discovery, Wnt signaling has had an association with cancer. When Wnt1 was discovered, it was first identified as a proto-oncogene in a mouse model for breast cancer. The fact that Wnt1 is a homolog of Wg shows that it is involved in embryonic development, which often calls for rapid cell division and migration. Misregulation of these processes can lead to tumor development via excess cell proliferation.[2]
Canonical Wnt activity pathway is involved in the development of benign and malignant breast tumors. Its presence is revealed by elevated levels of β-catenin in the nucleus and/or cytoplasm, which can be detected with immunohistochemical staining and Western blotting. Increased β-catenin expression is correlated with poor prognosis in breast cancer patients. This accumulation may be due to factors such as mutations in β-catenin, deficiencies in the β-catenin destruction complex, most frequently by mutations in structurally disordered regions of APC, overexpression of Wnt ligands, loss of inhibitors and/or decreased activity of regulatory pathways (such as the Wnt/calcium pathway).[26][41][42] Breast tumors can metastasize due to Wnt involvement in EMT. Research looking at metastasis of basal-like breast cancer to the lungs showed that repression of Wnt/β-catenin signaling can prevent EMT, which can inhibit metastasis.[43]
Wnt signaling was implicated in the development of other cancers. Changes in CTNNB1 expression, which is the gene that encodes β-catenin, can be measured in breast colorectal, melanoma, prostate, lung, and other cancers. Increased expression of Wnt ligand-proteins such as Wnt 1, Wnt2 and Wnt7A were observed in the development of glioblastoma, oesophageal cancer and ovarian cancer respectively. Other proteins that cause multiple cancer types in the absence of proper functioning include ROR1, ROR2, SFRP4, Wnt5A, WIF1 and those of the TCF/LEF family.[44]
The link between PGE2 and Wnt suggests that chronic inflammation-related increase of PGE2 may lead to activation of Wnt pathway in different tissues, resulting in carcinogenesis.[3]
Type II diabetes
Diabetes mellitus type 2 is a common disease that causes reduced insulin secretion and increased insulin resistance in the periphery. It results in increased blood glucose levels, or hyperglycemia, which can be fatal if untreated. Since Wnt signaling is involved in insulin sensitivity, malfunctioning of its pathway could be involved. Overexpression of Wnt5b, for instance, may increase susceptibility due to its role in adipogenesis, since obesity and type II diabetes have high comorbidity.[45] Wnt signaling is a strong activator of mitochondrial biogenesis. This leads to increased production of reactive oxygen species (ROS) known to cause DNA and cellular damage.[46] This ROS-induced damage is significant because it can cause acute hepatic insulin resistance, or injury-induced insulin resistance.[47] Mutations in Wnt signaling-associated transcription factors, such as TCF7L2, are linked to increased susceptibility.[48]
See also
References
- 1 2 Nusse R, Varmus HE (Jun 1992). "Wnt genes". Cell. 69 (7): 1073–87. doi:10.1016/0092-8674(92)90630-U. PMID 1617723.
- 1 2 3 4 5 6 Nusse R (Jan 2005). "Wnt signaling in disease and in development". Cell Research. 15 (1): 28–32. doi:10.1038/sj.cr.7290260. PMID 15686623.
- 1 2 3 Goessling W, North TE, Loewer S, Lord AM, Lee S, Stoick-Cooper CL, Weidinger G, Puder M, Daley GQ, Moon RT, Zon LI (Mar 2009). "Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration". Cell. 136 (6): 1136–47. doi:10.1016/j.cell.2009.01.015. PMC 2692708
 . PMID 19303855.
. PMID 19303855. - 1 2 Logan CY, Nusse R (2004). "The Wnt signaling pathway in development and disease". Annual Review of Cell and Developmental Biology. 20: 781–810. doi:10.1146/annurev.cellbio.20.010403.113126. PMID 15473860.
- 1 2 3 4 5 6 7 8 9 10 Komiya Y, Habas R (Apr 2008). "Wnt signal transduction pathways". Organogenesis. 4 (2): 68–75. doi:10.4161/org.4.2.5851. PMC 2634250. PMID 19279717.
- ↑ Nusse R, van Ooyen A, Cox D, Fung YK, Varmus H (1984). "Mode of proviral activation of a putative mammary oncogene (int-1) on mouse chromosome 15". Nature. 307 (5947): 131–6. Bibcode:1984Natur.307..131N. doi:10.1038/307131a0. PMID 6318122.
- ↑ Klaus A, Birchmeier W (May 2008). "Wnt signalling and its impact on development and cancer". Nature Reviews. Cancer. 8 (5): 387–98. doi:10.1038/nrc2389. PMID 18432252.
- ↑ Cadigan KM, Nusse R (Dec 1997). "Wnt signaling: a common theme in animal development". Genes & Development. 11 (24): 3286–305. doi:10.1101/gad.11.24.3286. PMID 9407023.
- ↑ Kurayoshi M, Yamamoto H, Izumi S, Kikuchi A (Mar 2007). "Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling". The Biochemical Journal. 402 (3): 515–23. doi:10.1042/BJ20061476. PMC 1863570. PMID 17117926.
- ↑ Nusse, Roel. "The Wnt Homepage". Retrieved 15 April 2013.
- 1 2 3 Rao TP, Kühl M (Jun 2010). "An updated overview on Wnt signaling pathways: a prelude for more". Circulation Research. 106 (12): 1798–806. doi:10.1161/CIRCRESAHA.110.219840. PMID 20576942.
- ↑ Schulte G, Bryja V (Oct 2007). "The Frizzled family of unconventional G-protein-coupled receptors". Trends in Pharmacological Sciences. 28 (10): 518–25. doi:10.1016/j.tips.2007.09.001. PMID 17884187.
- ↑ Habas R, Dawid IB (February 2005). "Dishevelled and Wnt signaling: is the nucleus the final frontier?". Journal of Biology. 4 (1): 2. doi:10.1186/jbiol22. PMC 551522. PMID 15720723.
- ↑ Minde DP, Anvarian Z, Rüdiger SG, Maurice MM (2011). "Messing up disorder: how do missense mutations in the tumor suppressor protein APC lead to cancer?" (PDF). Molecular Cancer. 10: 101. doi:10.1186/1476-4598-10-101. PMC 3170638. PMID 21859464.
- ↑ Minde, David P.; Radli, Martina; Forneris, Frederico; Maurice, Madelon M.; Rüdiger, Stefan G. D. (2013). Buckle, Ashley M, ed. "Large Extent of Disorder in Adenomatous Polyposis Coli Offers a Strategy to Guard Wnt Signalling against Point Mutations". PLoS ONE. 8 (10): e77257. doi:10.1371/journal.pone.0077257. PMC 3793970. PMID 24130866.
- 1 2 3 4 MacDonald BT, Tamai K, He X (Jul 2009). "Wnt/beta-catenin signaling: components, mechanisms, and diseases". Developmental Cell. 17 (1): 9–26. doi:10.1016/j.devcel.2009.06.016. PMC 2861485. PMID 19619488.
- ↑ Staal FJ, Clevers H (February 2000). "Tcf/Lef transcription factors during T-cell development: unique and overlapping functions". The Hematology Journal. 1 (1): 3–6. doi:10.1038/sj/thj/6200001 (inactive 2016-09-03). PMID 11920163.
- ↑ Gordon MD, Nusse R (Aug 2006). "Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors". The Journal of Biological Chemistry. 281 (32): 22429–33. doi:10.1074/jbc.R600015200. PMID 16793760.
- ↑ Sugimura R, Li L (Dec 2010). "Noncanonical Wnt signaling in vertebrate development, stem cells, and diseases". Birth Defects Research. Part C, Embryo Today. 90 (4): 243–56. doi:10.1002/bdrc.20195. PMID 21181886.
- 1 2 3 van Amerongen R, Nusse R (Oct 2009). "Towards an integrated view of Wnt signaling in development". Development. 136 (19): 3205–14. doi:10.1242/dev.033910. PMID 19736321.
- ↑ van Amerongen R, Fuerer C, Mizutani M, Nusse R (Sep 2012). "Wnt5a can both activate and repress Wnt/β-catenin signaling during mouse embryonic development". Developmental Biology. 369 (1): 101–14. doi:10.1016/j.ydbio.2012.06.020. PMC 3435145. PMID 22771246.
- ↑ Thrasivoulou C, Millar M, Ahmed A (Dec 2013). "Activation of intracellular calcium by multiple Wnt ligands and translocation of β-catenin into the nucleus: a convergent model of Wnt/Ca2+ and Wnt/β-catenin pathways". The Journal of Biological Chemistry. 288 (50): 35651–9. doi:10.1074/jbc.M112.437913. PMC 3861617. PMID 24158438.
- ↑ Inoki K, Ouyang H, Zhu T, Lindvall C, Wang Y, Zhang X, Yang Q, Bennett C, Harada Y, Stankunas K, Wang CY, He X, MacDougald OA, You M, Williams BO, Guan KL (Sep 2006). "TSC2 integrates Wnt and energy signals via a coordinated phosphorylation by AMPK and GSK3 to regulate cell growth". Cell. 126 (5): 955–68. doi:10.1016/j.cell.2006.06.055. PMID 16959574.
- ↑ Kuroda K, Kuang S, Taketo MM, Rudnicki MA (2013). "Canonical Wnt signaling induces BMP-4 to specify slow myofibrogenesis of fetal myoblasts". Skeletal Muscle. 3 (1): 5. doi:10.1186/2044-5040-3-5. PMC 3602004. PMID 23497616.
- ↑ Malinauskas T, Jones EY (Dec 2014). "Extracellular modulators of Wnt signalling". Current Opinion in Structural Biology. 29: 77–84. doi:10.1016/j.sbi.2014.10.003. PMID 25460271.
- 1 2 Minde DP, Radli M, Forneris F, Maurice MM, Rüdiger SG (2013). "Large extent of disorder in Adenomatous Polyposis Coli offers a strategy to guard Wnt signalling against point mutations". PLoS ONE. 8 (10): e77257. doi:10.1371/journal.pone.0077257. PMC 3793970. PMID 24130866.
- 1 2 3 4 Gilbert, Scott F. (2010). Developmental biology (9th ed.). Sunderland, Mass.: Sinauer Associates. ISBN 9780878933846.
- ↑ Vasiev, Bakhtier; Balter, Ariel; Chaplain, Mark; Glazier, James A.; Weijer, Cornelis J. (2010-05-11). "Modeling Gastrulation in the Chick Embryo: Formation of the Primitive Streak". PLoS ONE. 5 (5): e10571. Bibcode:2010PLoSO...510571V. doi:10.1371/journal.pone.0010571. PMC 2868022. PMID 20485500.
- ↑ Gilbert SF. Developmental Biology. 10th edition. Sunderland (MA): Sinauer Associates; 2014. Early Development in Birds. Print
- ↑ Ulloa F, Martí E (Jan 2010). "Wnt won the war: antagonistic role of Wnt over Shh controls dorso-ventral patterning of the vertebrate neural tube". Developmental Dynamics. 239 (1): 69–76. doi:10.1002/dvdy.22058. PMID 19681160.
- ↑ Zou Y (Sep 2004). "Wnt signaling in axon guidance". Trends in Neurosciences. 27 (9): 528–32. doi:10.1016/j.tins.2004.06.015. PMID 15331234.
- 1 2 Nusse R (May 2008). "Wnt signaling and stem cell control". Cell Research. 18 (5): 523–7. doi:10.1038/cr.2008.47. PMID 18392048.
- ↑ Bakre MM, Hoi A, Mong JC, Koh YY, Wong KY, Stanton LW (Oct 2007). "Generation of multipotential mesendodermal progenitors from mouse embryonic stem cells via sustained Wnt pathway activation". The Journal of Biological Chemistry. 282 (43): 31703–12. doi:10.1074/jbc.M704287200. PMID 17711862.
- ↑ Woll PS, Morris JK, Painschab MS, Marcus RK, Kohn AD, Biechele TL, Moon RT, Kaufman DS (Jan 2008). "Wnt signaling promotes hematoendothelial cell development from human embryonic stem cells". Blood. 111 (1): 122–31. doi:10.1182/blood-2007-04-084186. PMC 2200802. PMID 17875805.
- ↑ Kaldis P, Pagano M (Dec 2009). "Wnt signaling in mitosis". Developmental Cell. 17 (6): 749–50. doi:10.1016/j.devcel.2009.12.001. PMID 20059944.
- ↑ Willert K, Jones KA (Jun 2006). "Wnt signaling: is the party in the nucleus?". Genes & Development. 20 (11): 1394–404. doi:10.1101/gad.1424006. PMID 16751178.
- ↑ Hodge, Russ (2016-01-25). "Hacking the programs of cancer stem cells". medicalxpress.com. Medical Express. Retrieved 2016-02-12.
- ↑ Schambony A, Wedlich D. "Wnt Signaling and Cell Migration". Madame Curie Bioscience Database. Landes Bioscience. Retrieved 7 May 2013.
- ↑ Micalizzi DS, Farabaugh SM, Ford HL (Jun 2010). "Epithelial-mesenchymal transition in cancer: parallels between normal development and tumor progression". Journal of Mammary Gland Biology and Neoplasia. 15 (2): 117–34. doi:10.1007/s10911-010-9178-9. PMC 2886089. PMID 20490631.
- ↑ Abiola M, Favier M, Christodoulou-Vafeiadou E, Pichard AL, Martelly I, Guillet-Deniau I (Dec 30, 2009). "Activation of Wnt/beta-catenin signaling increases insulin sensitivity through a reciprocal regulation of Wnt10b and SREBP-1c in skeletal muscle cells". PLoS ONE. 4 (12): e8509. Bibcode:2009PLoSO...4.8509A. doi:10.1371/journal.pone.0008509. PMC 2794543. PMID 20041157.
- ↑ Howe LR, Brown AM (Jan 2004). "Wnt signaling and breast cancer". Cancer Biology & Therapy. 3 (1): 36–41. doi:10.4161/cbt.3.1.561. PMID 14739782.
- ↑ Taketo MM (Apr 2004). "Shutting down Wnt signal-activated cancer". Nature Genetics. 36 (4): 320–2. doi:10.1038/ng0404-320. PMID 15054482.
- ↑ DiMeo TA, Anderson K, Phadke P, Fan C, Feng C, Perou CM, Naber S, Kuperwasser C (Jul 2009). "A novel lung metastasis signature links Wnt signaling with cancer cell self-renewal and epithelial-mesenchymal transition in basal-like breast cancer". Cancer Research. 69 (13): 5364–73. doi:10.1158/0008-5472.CAN-08-4135. PMC 2782448. PMID 19549913.
- ↑ Anastas JN, Moon RT (Jan 2013). "WNT signalling pathways as therapeutic targets in cancer". Nature Reviews. Cancer. 13 (1): 11–26. doi:10.1038/nrc3419. PMID 23258168.
- ↑ Welters HJ, Kulkarni RN (Dec 2008). "Wnt signaling: relevance to beta-cell biology and diabetes". Trends in Endocrinology and Metabolism. 19 (10): 349–55. doi:10.1016/j.tem.2008.08.004. PMID 18926717.
- ↑ Yoon JC, Ng A, Kim BH, Bianco A, Xavier RJ, Elledge SJ (Jul 2010). "Wnt signaling regulates mitochondrial physiology and insulin sensitivity". Genes & Development. 24 (14): 1507–18. doi:10.1101/gad.1924910. PMC 2904941. PMID 20634317.
- ↑ Zhai L, Ballinger SW, Messina JL (Mar 2011). "Role of reactive oxygen species in injury-induced insulin resistance". Molecular Endocrinology. 25 (3): 492–502. doi:10.1210/me.2010-0224. PMC 3045736. PMID 21239612.
- ↑ Grant SF, Thorleifsson G, Reynisdottir I, Benediktsson R, Manolescu A, Sainz J, Helgason A, Stefansson H, Emilsson V, Helgadottir A, Styrkarsdottir U, Magnusson KP, Walters GB, Palsdottir E, Jonsdottir T, Gudmundsdottir T, Gylfason A, Saemundsdottir J, Wilensky RL, Reilly MP, Rader DJ, Bagger Y, Christiansen C, Gudnason V, Sigurdsson G, Thorsteinsdottir U, Gulcher JR, Kong A, Stefansson K (Mar 2006). "Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes". Nature Genetics. 38 (3): 320–3. doi:10.1038/ng1732. PMID 16415884.
External links
- Mouse Wnt proteins from Signaling Gateway Molecule Pages
- Dinasarapu AR, Saunders B, Ozerlat I, Azam K, Subramaniam S (Jun 2011). "Signaling gateway molecule pages--a data model perspective". Bioinformatics. 27 (12): 1736–8. doi:10.1093/bioinformatics/btr190. PMC 3106186. PMID 21505029.
- "Search Molecule Pages - Signaling Gateway".
- "The Wnt Homepage".
- "wntpathways".
- "KEGG PATHWAY: dme04310".
- mouse Wnt pathway from KEGG
- "KEGG PATHWAY: hsa04310".
- "KEGG PATHWAY: Wnt signaling pathway - Homo sapiens (human)".
- ABC News. "Can Grey Hair Be Reversed? Video - ABC News". ABC News.
- Netpath - A curated resource of signal transduction pathways in humans
- Wnt Proteins at the US National Library of Medicine Medical Subject Headings (MeSH)
- filip. "EPSF homepage".